La réplication de l’ADN est le moment où une cellule prépare sa descendance sans sacrifier son identité génétique. Derrière ce geste apparemment simple se cachent une ouverture contrôlée de la double hélice, une synthèse orientée, des mécanismes de correction et, à l’échelle de l’évolution, une source discrète mais décisive de diversité. Je vais ici montrer comment tout cela s’enchaîne, pourquoi le processus reste si fidèle et ce qu’il révèle sur l’histoire du vivant.
Ce qu’il faut retenir sur la copie de l’ADN
- Le mécanisme est semi-conservatif : chaque molécule fille garde un brin ancien et fabrique un brin neuf.
- L’ADN polymérase ne synthétise qu’en 5' vers 3' et a besoin d’une amorce pour démarrer.
- L’hélicase ouvre la double hélice, la primase pose l’amorce, la ligase soude les fragments, et les topoisomérases évitent la torsion excessive.
- La fidélité repose sur deux filets de sécurité : la relecture de la polymérase et la réparation des mésappariements.
- Les cellules eucaryotes utilisent plusieurs origines de réplication pour dupliquer rapidement de très grands chromosomes.
- Les erreurs qui échappent à la correction deviennent des mutations, donc une matière première pour l’évolution.
Ce que la cellule doit vraiment copier avant une division
Avant de se diviser, une cellule ne copie pas seulement quelques gènes utiles. Elle doit dupliquer tout son génome, c’est-à-dire l’ensemble des molécules d’ADN qui portent l’information héréditaire. Chez l’être humain, cela représente environ 3 milliards de paires de bases pour un génome haploïde, et près de 6 milliards dans la plupart des cellules somatiques.
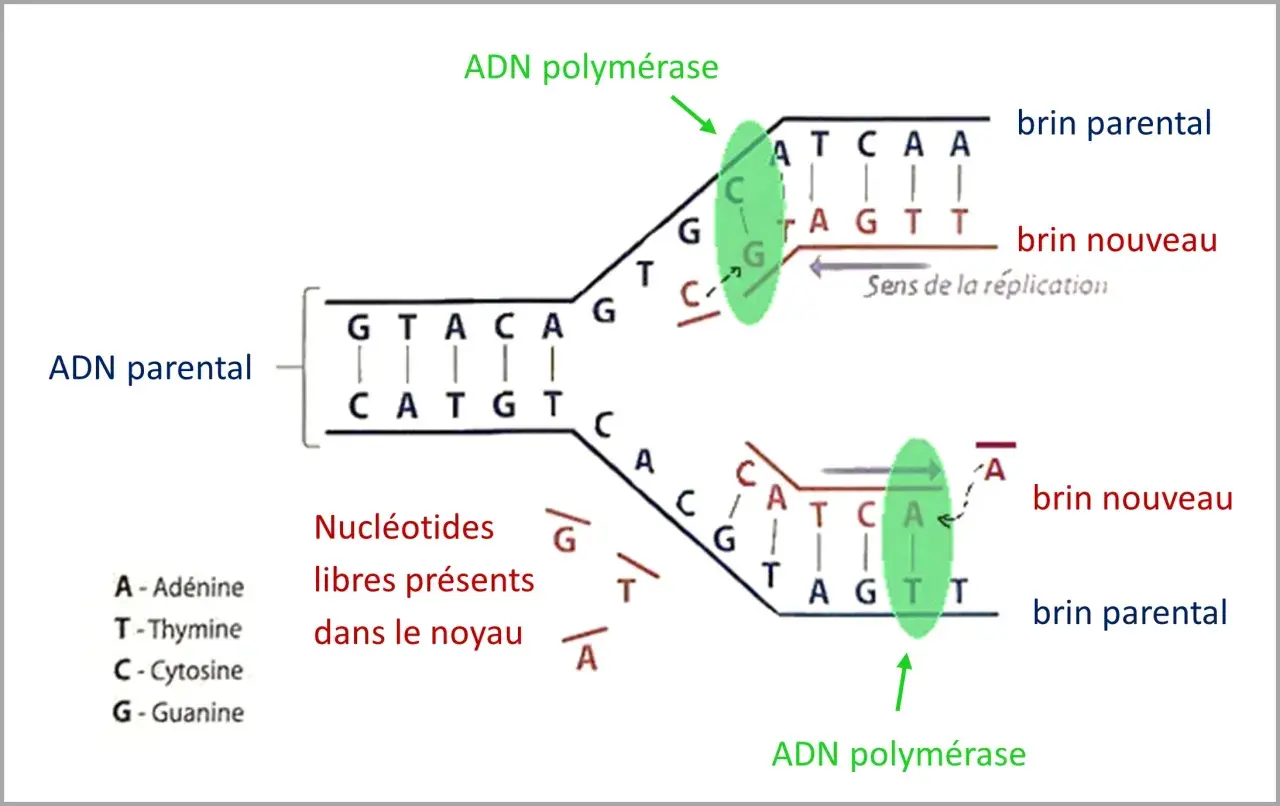
Je trouve utile de partir de là, parce qu’on comprend immédiatement pourquoi la précision est vitale. La molécule d’ADN est une double hélice dont les deux brins sont complémentaires : si l’un porte A, l’autre porte T ; si l’un porte G, l’autre porte C. La copie est dite semi-conservative, ce qui signifie qu’une nouvelle molécule conserve un brin ancien et synthétise un brin neuf en miroir.
Cette logique explique à la fois la stabilité des lignées cellulaires et la continuité du patrimoine génétique d’une génération à l’autre. Pour voir comment cette copie démarre vraiment, il faut maintenant entrer dans la fourche de réplication.
Comment la copie du génome se déroule pas à pas
L’initiation ouvre un site précis
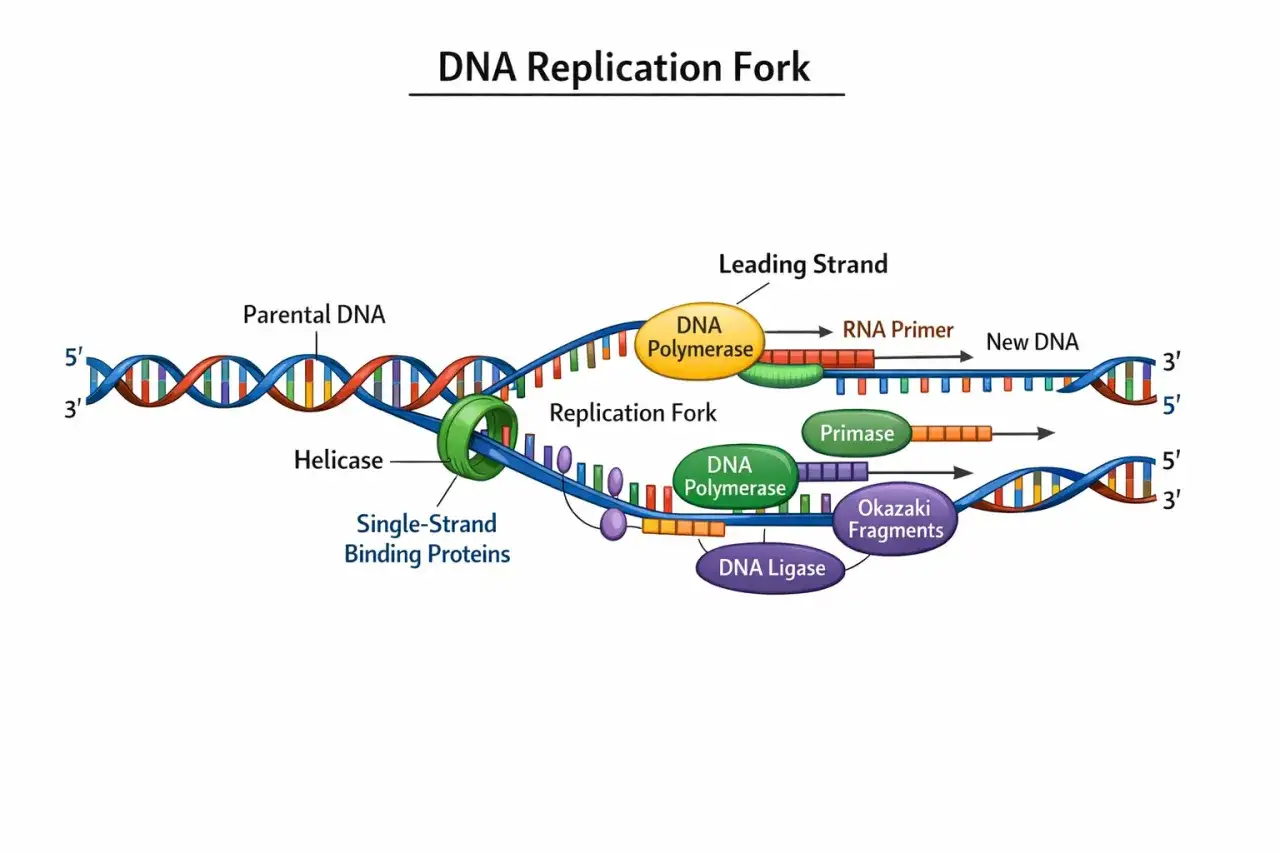
Tout commence sur une origine de réplication, une région où des protéines initiatrices recrutent la machinerie nécessaire. L’hélicase agit alors comme un moteur d’ouverture : elle sépare les deux brins en rompant les liaisons hydrogène qui maintiennent la double hélice. Cette zone active prend la forme d’une fourche de réplication, une structure en Y qui avance le long de la molécule.
À ce stade, les brins séparés sont fragiles et auraient tendance à se réassocier ou à former des structures instables. Des protéines de liaison à l’ADN simple brin les maintiennent donc ouverts et accessibles. C’est le point de départ du chantier.
L’élongation suit le sens 5' vers 3'
Je préfère toujours rappeler ce point, car il fait tomber beaucoup de confusions : l’ADN polymérase ne peut ajouter des nucléotides qu’à l’extrémité 3' du brin en cours de synthèse. Autrement dit, la chaîne nouvelle s’allonge uniquement dans le sens 5' vers 3'. Cette contrainte est centrale, parce qu’elle explique la différence entre le brin directeur, copié de façon continue, et le brin retardé, copié par fragments.
Sur le brin retardé, la cellule fabrique de courts segments appelés fragments d’Okazaki. Ils sont ensuite reliés entre eux pour former une séquence continue. Le point important n’est pas seulement la vitesse, mais la coordination : plusieurs enzymes doivent avancer ensemble sans se gêner, tout en respectant l’orientation des brins antiparallèles.
Lire aussi : Yaourt fermenté - Comprendre la vraie nature de ce produit
La terminaison ferme la copie
Quand deux fourches se rencontrent, la duplication s’achève. Chez les bactéries, les fourches finissent par se rejoindre sur un chromosome circulaire. Chez les eucaryotes, la fin est plus délicate, surtout aux extrémités des chromosomes linéaires, où interviennent des solutions spécifiques comme les télomères et, dans certaines cellules, la télomérase.
À partir de là, la vraie question devient presque mécanique : quelles protéines rendent ce ballet possible sans erreur majeure ? C’est ce que je détaille juste après.
Les enzymes qui rendent le processus possible
Je préfère lire cette machinerie comme une chaîne de travail, pas comme une simple liste d’enzymes. Chaque acteur prépare le suivant, et si l’un manque, toute la copie ralentit ou devient instable.
| Acteur | Rôle principal | Ce qu’il faut retenir |
|---|---|---|
| Hélicase | Ouvre la double hélice | Elle sépare les deux brins pour rendre la matrice accessible. |
| Primase | Fabrique une amorce ARN | L’ADN polymérase ne sait pas démarrer seule ; l’amorce lui donne un point de départ. |
| ADN polymérase | Ajoute les nucléotides complémentaires | Elle travaille en 5' vers 3' et corrige une partie de ses erreurs pendant la synthèse. |
| Protéines de liaison à l’ADN simple brin | Stabilisent les brins ouverts | Elles évitent que l’ADN ne se reforme trop tôt ou ne s’abîme. |
| Topoisomérase | Relâche la tension de l’ADN | Elle coupe et ressoude brièvement l’ADN pour empêcher le surenroulement. |
| Ligase | Soude les fragments | Elle referme les cassures entre les fragments d’Okazaki. |
| Télomérase | Protège les extrémités chromosomiques | Elle aide à résoudre le problème des chromosomes linéaires dans certaines cellules eucaryotes. |
Ce tableau montre bien une chose : la copie ne dépend pas d’une seule enzyme miracle, mais d’un ensemble très coordonné. C’est aussi pour cela qu’un défaut sur une seule étape peut avoir des conséquences visibles sur la stabilité du génome. Une fois ce décor posé, il reste à comprendre pourquoi le processus est si fidèle malgré tout.
Pourquoi la copie reste très fidèle sans être parfaite
La cellule doit gérer un volume d’information gigantesque, donc la fidélité n’est pas un luxe. Pourtant, aucune copie biologique n’est absolument parfaite. Les erreurs existent, mais elles sont contenues par plusieurs niveaux de contrôle. Le premier, c’est la relecture : la polymérase vérifie ce qu’elle vient d’ajouter et corrige une partie des mauvais appariements. Le second, c’est la réparation des mésappariements, qui intervient après coup pour rectifier les fautes passées entre les mailles du filet.
En pratique, cela réduit fortement le nombre d’erreurs. La relecture améliore déjà la fidélité d’un ordre de grandeur important, et les systèmes de réparation abaissent encore le risque final. Je vois souvent une erreur d’interprétation chez les débutants : ils imaginent qu’une mutation visible doit forcément apparaître à chaque copie. En réalité, la plupart des fautes sont réparées, et beaucoup de celles qui subsistent n’ont aucun effet détectable.
- Substitution : un nucléotide est remplacé par un autre.
- Insertion : un ou plusieurs nucléotides s’ajoutent.
- Délétion : un ou plusieurs nucléotides disparaissent.
Les erreurs ne viennent d’ailleurs pas uniquement de la copie elle-même. Elles peuvent aussi résulter de dommages chimiques ou physiques, comme les rayonnements ou certains agents mutagènes. Ce mélange entre exactitude et imperfection explique pourquoi la réplication stabilise le vivant tout en laissant subsister une marge d’innovation. Et c’est précisément ce qui devient intéressant quand on compare les grands groupes d’organismes.
Procaryotes et eucaryotes ne répliquent pas leur ADN au même rythme
La logique générale reste la même, mais l’organisation change beaucoup selon la taille du génome et la structure des chromosomes. Chez les bactéries, le chromosome est souvent circulaire et la duplication part d’une seule origine principale. Chez les eucaryotes, les chromosomes sont linéaires, plus longs et plus nombreux, ce qui impose plusieurs origines réparties sur chaque chromosome pour finir la copie dans un temps raisonnable.
| Critère | Procaryotes | Eucaryotes |
|---|---|---|
| Structure du génome | Souvent un chromosome circulaire | Chromosomes linéaires organisés en chromatine |
| Origines de réplication | Généralement une origine principale | Plusieurs origines par chromosome |
| Vitesse et organisation | Un génome d’E. coli d’environ 4,6 millions de paires de bases peut être dupliqué en environ 40 minutes, avec des fourches qui avancent à 500 à 1000 nucléotides par seconde | La vitesse par fourche est plus modérée, mais la multiplication des origines compense la taille du génome |
| Moment de la copie | Plus souple selon l’état physiologique de la cellule | Strictement limité à la phase S du cycle cellulaire |
| Extrémités des chromosomes | Pas de vrai problème d’extrémité sur un chromosome circulaire | Nécessite des solutions spécifiques, notamment les télomères |
Un détail compte particulièrement chez les eucaryotes : les origines doivent être « licenciées », c’est-à-dire préparées de manière contrôlée pour ne s’activer qu’une seule fois par cycle. Cette précaution évite qu’une même région soit copiée deux fois. À mon sens, c’est l’un des meilleurs exemples de régulation fine en biologie cellulaire, et il ouvre naturellement sur la place de ce mécanisme dans l’évolution.
Pourquoi une copie fidèle n’empêche pas l’évolution
La réplication de l’ADN ne sert pas seulement à conserver l’identité d’une cellule ; elle produit aussi, à faible dose, de la variation. Les erreurs qui échappent à la correction deviennent des mutations, et ce sont elles qui alimentent la diversité génétique. Selon leur emplacement et leur nature, elles peuvent être neutres, défavorables ou parfois avantageuses dans un environnement donné.
C’est là que la biologie évolutive prend tout son sens. Une population ne change pas parce que chaque copie est mauvaise, mais parce que des variantes apparaissent de manière ponctuelle, puis sont triées par la sélection naturelle, la dérive génétique ou les contraintes du milieu. La copie fidèle assure la continuité ; l’imperfection résiduelle fournit la matière première du changement. Je trouve que c’est l’un des plus beaux compromis du vivant.Les travaux d’histoire du vivant suggèrent aussi que les systèmes de réplication de l’ADN sont apparus tardivement, après des formes de chimie biologique plus anciennes dominées par l’ARN et les protéines. Autrement dit, la grande stabilité du génome n’est pas un point de départ absolu : c’est le résultat d’une longue évolution des machineries cellulaires. Si je devais résumer l’idée essentielle, je dirais que la réplication de l’ADN protège l’héritage biologique tout en laissant juste assez d’écart pour que l’évolution puisse travailler.