Un schéma crossing over bien construit permet de comprendre d’un seul coup d’œil comment deux chromosomes homologues échangent des fragments d’ADN pendant la méiose. Dans cet article, je montre comment lire ce type de dessin, quels éléments il faut repérer et pourquoi ce mécanisme compte autant en biologie de l’évolution. L’enjeu est simple : distinguer un vrai échange génétique d’un croisement graphique mal interprété.
Les repères essentiels avant de lire le schéma
- Le crossing-over a lieu pendant la prophase I de la méiose.
- Il se produit entre chromatides non sœurs de chromosomes homologues.
- Le résultat est un chromosome recombinant, pas un changement du nombre de chromosomes.
- Le mécanisme augmente la diversité des gamètes et nourrit la variation sur laquelle agit la sélection naturelle.
- Un échange inégal peut créer des duplications ou des délétions de segments d’ADN.
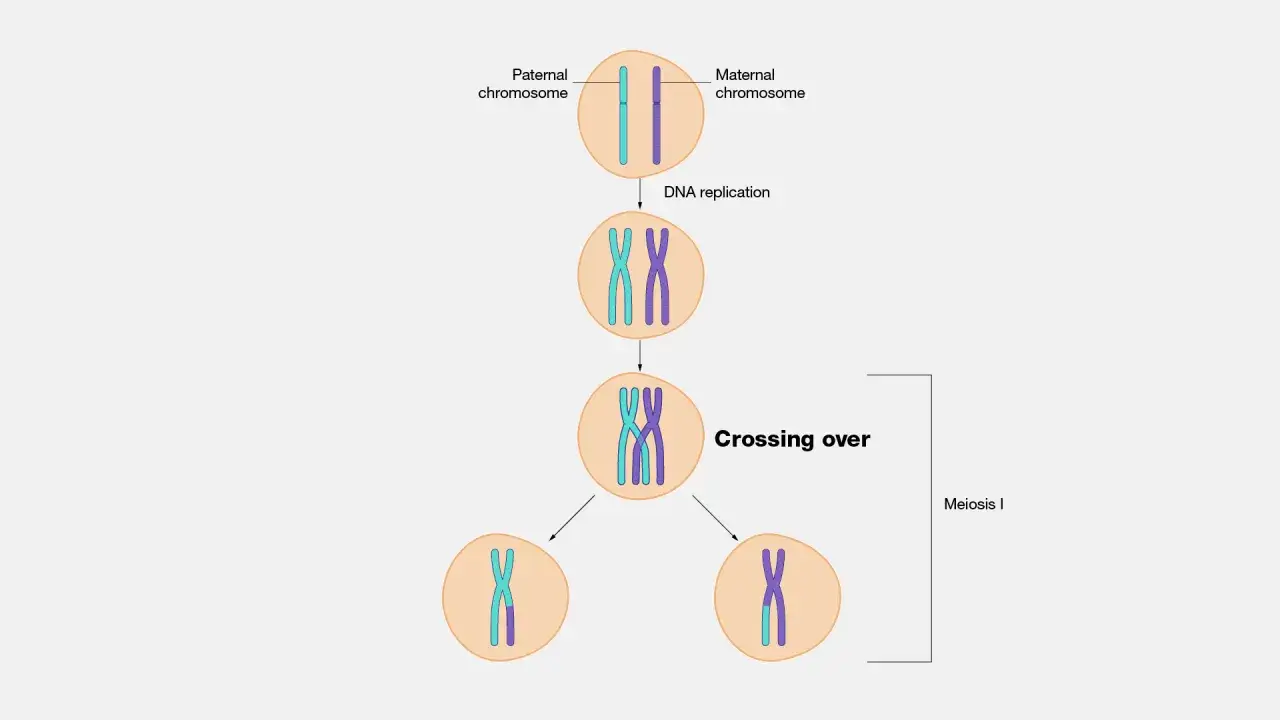
Ce qu’on voit vraiment dans un schéma du crossing-over
Je commence toujours par trois repères visuels : la paire de chromosomes homologues, les chromatides non sœurs et le point de contact qui marque l’échange. Le terme locus désigne simplement l’emplacement d’un gène sur un chromosome, et c’est utile pour comprendre pourquoi deux segments échangés ne racontent pas la même histoire génétique. Dans beaucoup de schémas scolaires, les couleurs servent seulement à suivre l’origine parentale des fragments ; elles ne veulent pas dire que les chromosomes sont réellement rouges ou bleus.
Autrement dit, le dessin simplifie la réalité, mais il ne doit jamais faire perdre de vue la logique biologique. Voici les éléments que je recommande de lire en priorité :
| Élément | Ce qu’il représente | Ce qu’il faut retenir |
|---|---|---|
| Chromosomes homologues | Les deux chromosomes d’une même paire, l’un d’origine maternelle, l’autre paternelle | Ils portent les mêmes gènes aux mêmes loci, mais pas forcément les mêmes allèles |
| Chromatides non sœurs | Les chromatides appartenant à deux homologues différents | L’échange se fait entre elles, pas entre chromatides sœurs |
| Bivalent ou tétrade | L’ensemble formé par les deux homologues appariés, soit quatre chromatides | C’est la structure clé visible pendant la prophase I |
| Complexe synaptonémal | Une structure protéique qui stabilise l’alignement des homologues | Elle aide à organiser l’appariement avant l’échange |
| Chiasma | Le point de croisement visible après l’échange | Ce n’est pas un simple trait graphique, c’est la trace d’une recombinaison |
| Chromosome recombinant | Le chromosome portant un nouveau mélange de fragments parentaux | Le matériel génétique est réassocié, pas inventé de zéro |
Une fois ces repères posés, le déroulement devient beaucoup plus lisible. Et c’est justement ce déroulement qu’il faut savoir raconter sans hésiter.
Le déroulement exact de l’échange pendant la méiose
Le schéma pédagogique condense un processus cellulaire précis, mais la chronologie reste la même. Je le reformule souvent en cinq temps parce que cela évite les confusions :
- Appariement des homologues : les deux chromosomes d’une même paire se rapprochent et s’alignent.
- Formation du bivalent : les quatre chromatides sont regroupées dans une structure commune.
- Échange de segments : des fragments d’ADN sont transférés entre chromatides non sœurs.
- Résolution du chiasma : la jonction est tranchée et stabilisée, ce qui fixe le nouvel agencement.
- Séparation méiotique : les homologues se séparent en méiose I, puis les chromatides en méiose II.
Le point le plus important, c’est que l’échange intervient avant la séparation des chromosomes homologues, donc pendant la prophase I. Je vois souvent des schémas trop simplifiés qui donnent l’impression d’un simple « croisement » de lignes ; en réalité, la cellule orchestre un échange de matériel génétique beaucoup plus structuré. Si ce moment du cycle est clair, le reste du mécanisme devient intuitif.
Pourquoi ce mécanisme alimente la diversité et l’évolution
Le crossing-over ne crée pas une nouvelle information à partir de rien ; il recompose l’ADN déjà présent en nouvelles associations d’allèles. C’est précisément ce type de variation qui permet à une population de ne pas répondre tous ses individus de la même façon à un changement de milieu, à un pathogène ou à une pression de sélection.
Chez l’humain, le seul brassage interchromosomique peut déjà produire 223, soit 8 388 608 combinaisons de chromosomes parentaux dans les gamètes. Le crossing-over ajoute une couche supplémentaire, parce qu’il réorganise les segments à l’intérieur même d’un chromosome. Je trouve utile de séparer ces deux niveaux, car ils sont souvent confondus alors qu’ils ne mélangent pas le vivant de la même manière.
| Source de diversité | Ce qu’elle mélange | Repère chiffré | Intérêt évolutif |
|---|---|---|---|
| Brassage interchromosomique | Les chromosomes entiers | 223 chez l’humain, soit environ 8,4 millions de combinaisons | Répartition aléatoire des paires de chromosomes pendant la méiose I |
| Crossing-over | Des segments d’un même chromosome | 1 centimorgan (cM) correspond à environ 1 % de descendants recombinés | Création de nouvelles combinaisons d’allèles sur un même chromosome |
Cette fréquence de recombinaison sert aussi en cartographie génétique : plus deux loci sont éloignés, plus il y a de chances qu’un échange les sépare. En pratique, le centimorgan mesure donc une fréquence de recombinaison, pas une distance physique stricte. C’est un détail technique, mais il explique pourquoi le crossing-over est autant un sujet de génétique qu’un sujet d’évolution.
Mais pour bien lire un schéma, il faut aussi savoir éviter les erreurs qui reviennent sans cesse.
Les pièges de lecture que je vois le plus souvent
- Confondre chromosomes homologues et chromatides sœurs : l’échange n’a pas lieu entre les deux copies d’un même chromosome, mais entre les chromatides de deux homologues différents.
- Prendre le crossing-over pour une mutation : le mécanisme réorganise des segments déjà présents ; il ne modifie pas forcément la séquence de base.
- Le placer au mauvais moment : il se déroule pendant la prophase I, pas après la séparation des chromosomes homologues.
- Imaginer que toutes les chromatides changent : en général, seules certaines chromatides portent les fragments échangés.
- Lire le chiasma comme une simple décoration graphique : c’est un indice cytologique réel, pas une ligne de style.
- Oublier que la couleur des chromosomes est conventionnelle : dans les schémas, elle sert à suivre l’origine parentale, pas la matière réelle.
Quand je corrige des copies, je regarde surtout trois mots : homologue, non sœur et prophase I. S’ils sont justes, le raisonnement tient généralement. Et il reste enfin un cas particulier qui mérite une section à part : l’échange inégal.
Quand l’échange devient inégal
C’est le versant moins « propre » du phénomène. Quand les homologues s’alignent mal, l’échange porte sur des segments décalés et peut produire un chromosome avec un fragment en trop, l’autre avec un fragment en moins. On parle alors de crossing-over inégal.
| Aspect | Crossing-over classique | Crossing-over inégal |
|---|---|---|
| Alignement | Précis | Décalé |
| Résultat immédiat | Échange équilibré de fragments | Duplication sur un chromosome, délétion sur l’autre |
| Effet évolutif | Nouvelle combinaison d’allèles | Apparition possible de copies supplémentaires de gènes |
| Risque biologique | Faible si le mécanisme est bien régulé | Peut favoriser des déséquilibres ou des maladies génétiques |
Je trouve ce point essentiel pour la biologie de l’évolution, parce qu’une duplication peut offrir à une copie de gène le temps de diverger. Autrement dit, une cellule peut perdre de la stabilité à court terme, mais une lignée gagner une matière première évolutive à long terme. C’est ainsi que se forment et s’enrichissent certaines familles multigéniques, c’est-à-dire des groupes de gènes apparentés issus de duplications successives.
Lire correctement le schéma en examen et retenir l’essentiel
- Repère d’abord la paire de chromosomes homologues.
- Vérifie que l’échange se fait entre chromatides non sœurs.
- Replace l’événement en prophase I, au moment de l’appariement.
- Observe le résultat final : des chromosomes recombinés, pas une cellule avec plus ou moins de chromosomes.
Si je devais résumer la lecture en une seule méthode, je dirais ceci : un bon dessin doit montrer le contexte méiotique, le point d’échange et la conséquence génétique. Quand ces trois éléments sont clairs, on ne confond plus un simple croisement graphique avec un vrai mécanisme de recombinaison, et le schéma devient un outil solide pour comprendre la diversité du vivant.