La loi de Hardy-Weinberg sert de repère central en génétique des populations : elle décrit ce qui se passe quand les fréquences alléliques et génotypiques restent stables d’une génération à l’autre. Je vais vous montrer comment lire l’équation, dans quelles conditions elle fonctionne vraiment, et surtout comment interpréter un écart sans tomber dans les contresens classiques. C’est un modèle simple en apparence, mais il devient très parlant dès qu’on le relie à l’évolution, à la sélection et aux données de terrain.
Ce qu’il faut garder en tête
- Le modèle sert de référence théorique, pas de description parfaite du vivant.
- Avec deux allèles, on utilise p + q = 1 et p² + 2pq + q² = 1.
- Il suppose une grande population, des accouplements aléatoires et l’absence de mutation, de migration et de sélection.
- Un écart peut signaler une force évolutive, mais aussi un biais d’échantillonnage ou de génotypage.
- On l’emploie pour estimer des fréquences attendues, contrôler des données et repérer des signaux d’évolution.
Ce que décrit vraiment ce modèle
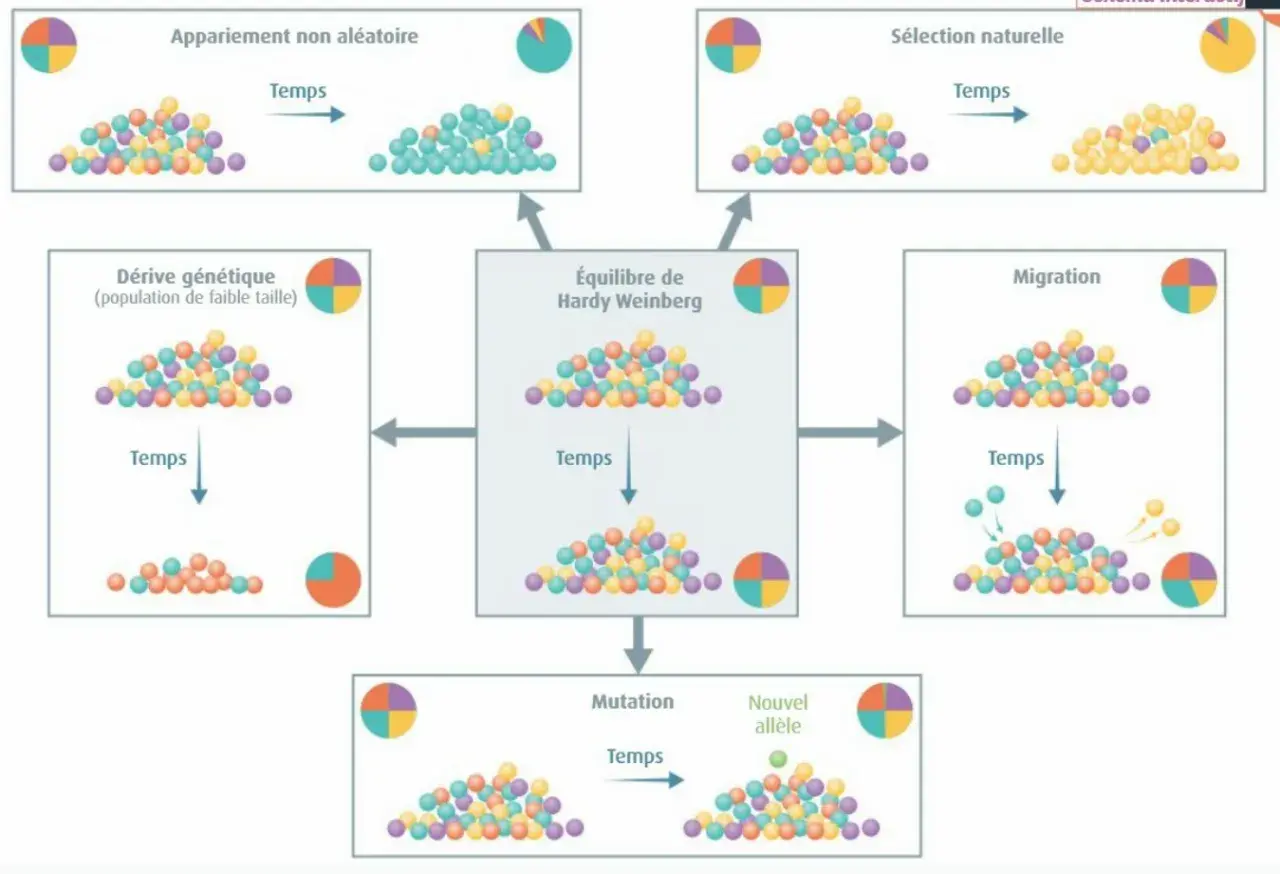
Je le présente souvent comme un modèle nul : tant qu’aucune force évolutive importante ne vient perturber la population, la diversité génétique attendue ne bouge pas. On parle surtout des fréquences alléliques et des fréquences génotypiques, deux notions qu’on confond encore trop souvent. L’idée n’est pas de dire que l’évolution s’arrête, mais de disposer d’un point zéro pour savoir si quelque chose pousse la population à changer.
Dans une population idéale, les unions sont aléatoires, l’effectif est grand, et les allèles ne donnent ni avantage ni désavantage particulier. Dès qu’on ajoute un facteur comme la sélection, la migration ou la dérive génétique, on ne regarde plus une attente théorique mais une population en mouvement. Pour passer de cette idée générale aux calculs concrets, il faut regarder l’équation elle-même.
Lire l’équation sans se perdre
Pour deux allèles seulement, la logique est simple : si la fréquence de A vaut p et celle de a vaut q, alors p + q = 1. Les génotypes attendus se répartissent selon p² pour AA, 2pq pour Aa et q² pour aa. Ce qui compte, c’est que la somme fasse 1, mais surtout que le terme 2pq rappelle à quel point les hétérozygotes peuvent être nombreux même quand un allèle est rare.
Un exemple chiffré utile en pratique
| Fréquence de l’allèle a | Fréquence de l’allèle A | AA | Aa | aa | Lecture rapide |
|---|---|---|---|---|---|
| 0,10 | 0,90 | 81 % | 18 % | 1 % | Beaucoup de porteurs, peu d’homozygotes récessifs |
| 0,01 | 0,99 | 98,01 % | 1,98 % | 0,01 % | Les atteints deviennent très rares, mais les porteurs restent présents |
Ce petit tableau vaut mieux qu’une longue explication abstraite. Il montre pourquoi un allèle pathogène peut rester discret dans les diagnostics tout en circulant largement dans la population. Et si le problème porte sur plus de deux allèles, le raisonnement reste le même, même si la formule se généralise.
La suite logique consiste donc à demander dans quelles conditions ces chiffres ont réellement un sens biologique.
Les conditions à respecter pour qu’il tienne
Le modèle repose sur quelques hypothèses strictes. Je préfère les lire comme des garde-fous : plus elles sont respectées, plus la comparaison entre l’attendu et l’observé est solide. En pratique, la population doit être suffisamment grande, les accouplements doivent être aléatoires pour le locus étudié, et il ne doit pas y avoir de mutation, de migration ni de sélection notable.
| Condition | Ce qu’elle implique concrètement | Si elle manque |
|---|---|---|
| Grande population | Les effets du hasard restent limités | La dérive génétique peut faire varier les fréquences par simple chance |
| Accouplements aléatoires | Les génotypes se forment sans préférence particulière | On obtient des excès ou des manques d’hétérozygotes ou d’homozygotes |
| Pas de sélection | Aucun génotype n’a d’avantage reproductif durable | Les fréquences changent dans une direction biologique identifiable |
| Pas de mutation | Aucun nouvel allèle n’apparaît pendant l’intervalle étudié | La diversité allélique se renouvelle |
| Pas de migration | La population reste génétiquement fermée | Un flux de gènes introduit ou retire des allèles |
Il faut ajouter une nuance que beaucoup d’étudiants oublient : le modèle est mathématiquement formulé pour une population infinie, mais en biologie on cherche surtout une population assez grande pour que la dérive soit négligeable. Autre point de vigilance, les loci liés au sexe demandent davantage de prudence que les loci autosomiques. Dès qu’une de ces hypothèses saute, l’écart devient intéressant en soi.
Et c’est justement cet écart qui donne au modèle sa vraie valeur d’outil d’analyse.
Pourquoi les populations réelles s’en écartent
Je me méfie toujours d’une lecture trop rapide : un écart au modèle ne dit pas immédiatement quelle force évolutive agit. Il faut d’abord distinguer les causes biologiques des causes techniques. Une sélection naturelle peut favoriser un allèle, mais un petit effectif peut aussi suffire à produire une variation aléatoire marquée, sans direction particulière.- Sélection naturelle : certains génotypes laissent plus de descendants que d’autres.
- Dérive génétique : dans une petite population, le hasard change les fréquences d’une génération à l’autre.
- Flux de gènes : des individus arrivent ou partent, et modifient les fréquences alléliques.
- Accouplements non aléatoires : la consanguinité ou le choix du partenaire modifient la répartition des génotypes.
- Biais de mesure : un échantillon mal conçu ou un génotypage imparfait peut fabriquer un faux signal.
Lire aussi : Acides aminés essentiels - Le guide complet pour votre santé
Les erreurs que je vois le plus souvent
La première erreur consiste à confondre fréquence allélique et fréquence génotypique. La deuxième est de conclure trop vite à une sélection alors qu’un effet de structure de population suffit parfois à expliquer l’écart. La troisième, plus discrète, consiste à ignorer la taille de l’échantillon : avec peu d’individus, les résultats sont facilement instables.
En clair, l’écart n’est pas un verdict, c’est un indice. C’est précisément pour cela que ce modèle reste si utile en génétique des populations.
Comment on l’utilise en génétique et en évolution
Dans la pratique, j’y vois trois usages majeurs. D’abord, il permet d’estimer les fréquences attendues quand on connaît la fréquence d’un allèle. Ensuite, il sert de test de cohérence : si les génotypes observés s’éloignent trop de l’attendu, il faut chercher une explication. Enfin, il aide à lire l’évolution elle-même, parce qu’un écart stable dans le temps peut révéler une pression de sélection, une migration ou une structure de population.
Il est aussi très parlant en génétique médicale. Si un allèle récessif pathogène a une fréquence de 1 %, on attend environ 1,98 % de porteurs sains et seulement 0,01 % d’homozygotes atteints dans une population idéale. Ce type de calcul reste une approximation, mais il donne tout de suite un ordre de grandeur utile pour raisonner sur le dépistage ou sur la rareté apparente d’une maladie.
Dans les études de conservation, l’idée est similaire : un excès d’homozygotes ou une variation inattendue des fréquences peut alerter sur un petit effectif, un isolement géographique ou une reproduction non aléatoire. Autrement dit, le modèle ne sert pas seulement à répondre à un exercice, il sert à lire une situation biologique réelle.
Reste à savoir comment interpréter proprement ce que l’on observe, sans surjouer la certitude.
Quand le modèle vous aide à interpréter un résultat réel
Si je devais résumer l’approche à garder, je dirais ceci : commencez par calculer l’attendu, comparez-le à l’observé, puis cherchez seulement ensuite la cause de l’écart. Cette méthode évite deux pièges très fréquents, à savoir attribuer un résultat au hasard sans vérifier les chiffres, ou au contraire voir une sélection partout. En biologie, la discipline de lecture compte autant que la formule.
- Comparez toujours des effectifs ou des fréquences calculées avec soin, pas seulement une intuition visuelle.
- Vérifiez si le locus étudié est autosomique, lié au sexe, ou soumis à une structure particulière de population.
- Ne confondez pas un déséquilibre génotypique avec une preuve directe d’évolution.
- Contrôlez l’échantillonnage et la qualité des données avant d’interpréter le signal biologique.
- Gardez en tête qu’un bon modèle simplifie la réalité, mais ne l’annule jamais.
Au fond, l’intérêt de la loi de Hardy-Weinberg est là : elle donne un repère stable pour comprendre pourquoi une population reste proche d’un équilibre, ou au contraire pourquoi elle s’en éloigne. C’est ce qui en fait un outil aussi utile en cours qu’en recherche. Si vous retenez une seule idée, gardez celle-ci : un bon calcul n’explique pas tout, mais il empêche de raconter n’importe quoi à partir d’un simple écart.