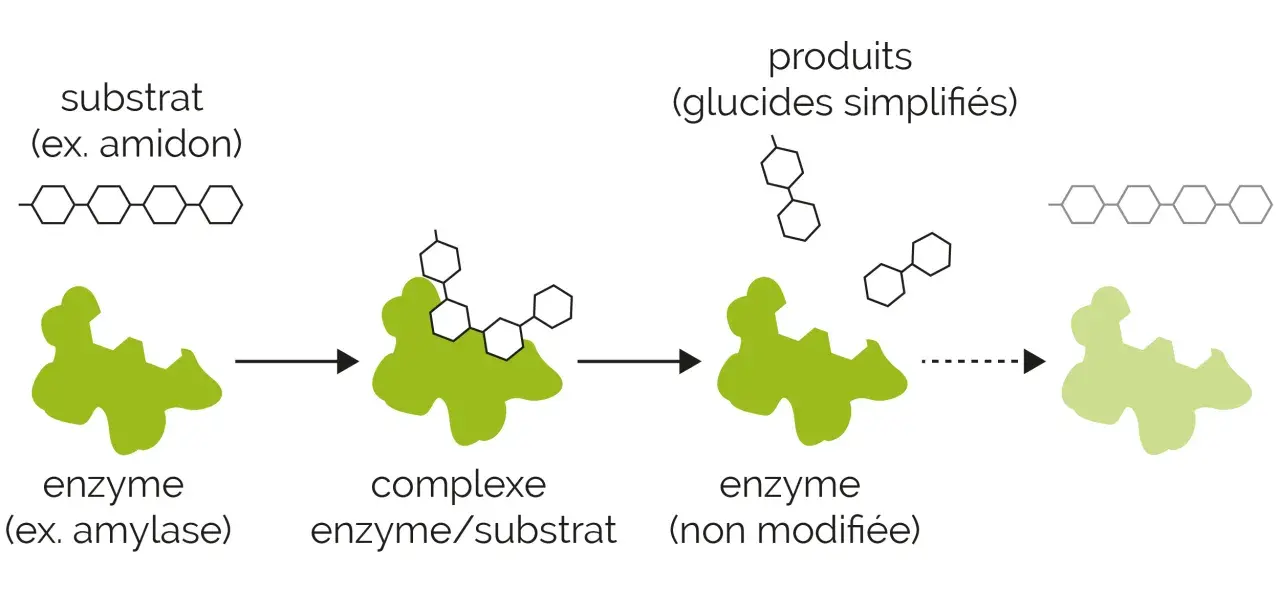
Le complexe enzyme-substrat est l’étape la plus discrète, mais aussi la plus décisive, de nombreuses réactions biologiques. C’est à ce moment précis qu’une enzyme reconnaît sa molécule cible, la maintient dans la bonne orientation puis facilite sa transformation en produit. Dans cet article, j’explique comment ce complexe se forme, pourquoi il accélère les réactions et ce qu’il révèle sur l’évolution des enzymes.
L’essentiel à retenir sur le complexe enzyme-substrat
- Il s’agit d’une structure transitoire formée quand une enzyme se lie à son substrat.
- Le site actif apporte la spécificité, mais il n’est pas toujours rigide.
- Le complexe abaisse surtout l’énergie d’activation, sans modifier le bilan final de la réaction.
- La température, le pH, les inhibiteurs et les mutations peuvent le stabiliser ou le déstabiliser.
- L’évolution des enzymes repose souvent sur des ajustements fins de cette reconnaissance moléculaire.
Ce que désigne vraiment le complexe enzyme-substrat
Une enzyme n’agit pas à distance : elle doit d’abord rencontrer son substrat et se fixer sur lui dans une zone précise appelée site actif. Cette association forme un complexe temporaire, réversible, qui existe juste assez longtemps pour permettre la réaction chimique. Une fois les produits formés, l’enzyme est libérée et peut recommencer un nouveau cycle.
Je trouve utile de distinguer ce point dès le départ, parce qu’on imagine parfois une liaison stable, presque définitive. En réalité, le complexe enzyme-substrat est une rencontre brève, mais extraordinairement efficace. Quand une réaction implique deux substrats, on parle parfois de complexe ternaire ; quand il n’y en a qu’un, on parle de complexe binaire.
La vraie question est donc simple : comment une interaction aussi courte peut-elle transformer une réaction lente en réaction rapide ?
Comment le complexe se forme et ce qu’il change dans la réaction
Le mécanisme se comprend bien si l’on suit la séquence dans l’ordre. Le substrat diffuse dans le milieu, entre dans le site actif, s’y stabilise un instant, puis la chimie proprement dite démarre. L’enzyme ne fabrique pas le produit à la place du substrat ; elle crée surtout un environnement favorable pour que la transformation devienne beaucoup plus facile.
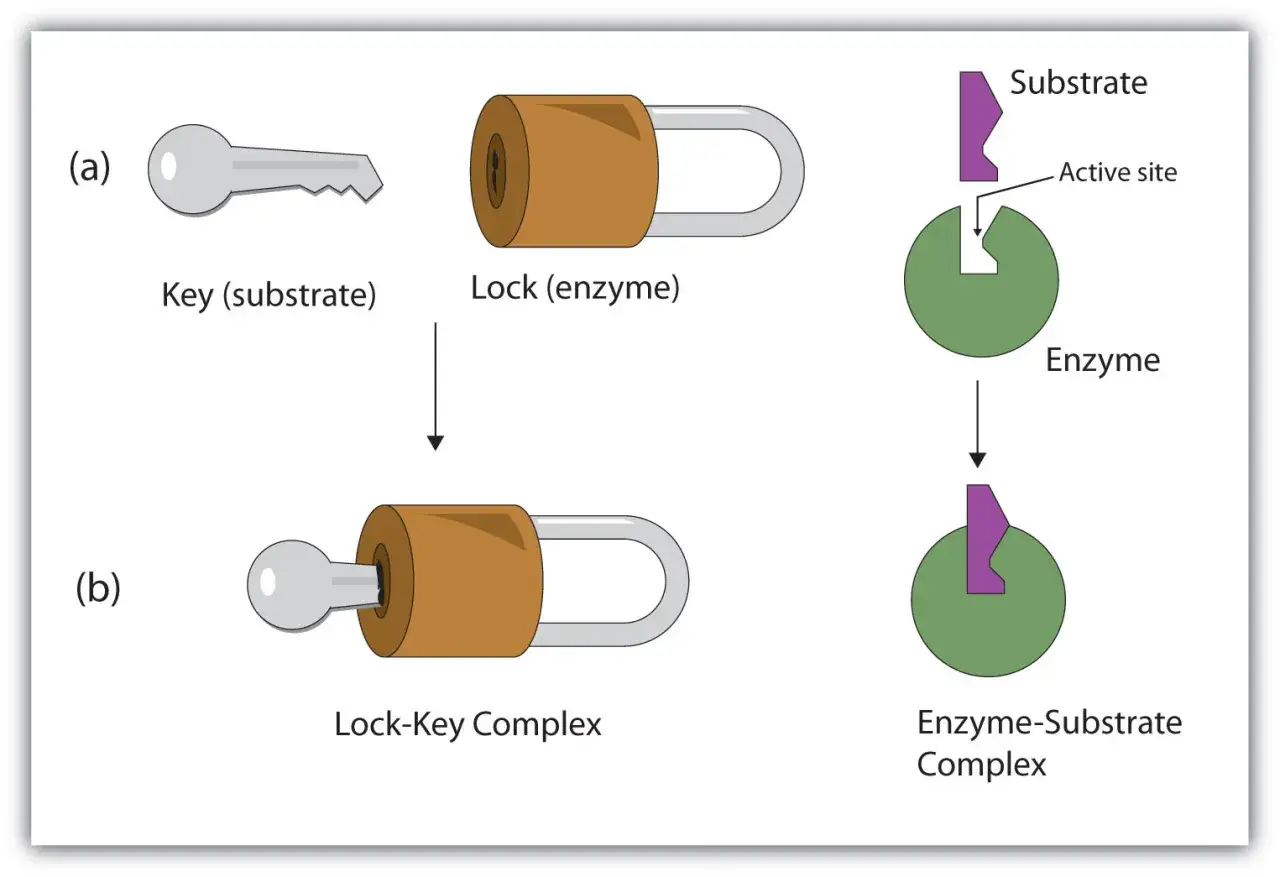
La logique clé-serrure et sa limite
Le modèle de la clé et de la serrure reste utile pour visualiser la complémentarité entre l’enzyme et son substrat. Il rappelle que la forme et la chimie du site actif doivent correspondre à la molécule ciblée. Mais ce modèle est trop rigide si on le prend au pied de la lettre.
Le modèle de l’ajustement induit est plus proche de ce qu’on observe souvent : la liaison du substrat provoque une légère modification de la forme de l’enzyme, et parfois du substrat lui-même. Autrement dit, le site actif n’est pas un moule figé. Il bouge juste assez pour mieux accueillir la molécule et orienter les groupes chimiques au bon endroit.
La stabilisation de l’état de transition
Je précise un détail souvent mal compris : une enzyme n’accélère pas une réaction en “ajoutant de l’énergie”. Elle l’accélère en abaissant l’énergie d’activation, c’est-à-dire la barrière à franchir pour passer des réactifs aux produits. L’enzyme y parvient en stabilisant l’état de transition, en rapprochant les bonnes parties du substrat et en limitant les mouvements inutiles.
C’est là que le complexe devient vraiment intéressant. Au lieu de laisser les molécules se rencontrer au hasard, l’enzyme les place dans une géométrie favorable. Elle peut aussi exclure l’eau, modifier localement les charges ou tendre certaines liaisons pour rendre la rupture ou la formation de liaison plus facile. Certaines enzymes très actives peuvent ainsi enchaîner de l’ordre de 10^3 conversions par seconde, même si cette valeur varie énormément selon l’enzyme et le substrat.
La libération du produit
Une fois la réaction accomplie, les produits n’ont souvent plus la même affinité pour le site actif. Ils se détachent, ce qui libère l’enzyme pour un nouveau cycle catalytique. Cette étape paraît secondaire, mais elle est essentielle : si le produit restait trop longtemps accroché, l’enzyme serait vite “bloquée”.
À ce stade, les limites du système apparaissent surtout quand le milieu change, et c’est précisément ce qui rend la suite si importante.
Les conditions qui renforcent ou fragilisent la liaison
Le complexe ne se forme pas dans le vide. Il dépend de la température, du pH, de la concentration en substrat, de la présence de cofacteurs et de l’état structural de l’enzyme. Quand l’une de ces variables sort de la zone de confort de la protéine, l’affinité baisse, la catalyse ralentit ou la structure se dégrade.
Le plus souvent, les débutants sous-estiment un point simple : plus de substrat n’accélère pas toujours la réaction. Au début, la vitesse augmente parce que davantage de sites actifs sont occupés. Mais quand les sites actifs sont saturés, ajouter encore du substrat ne change presque plus rien.
| Facteur | Effet sur le complexe | Ce que cela signifie en pratique |
|---|---|---|
| pH | Il modifie les charges des acides aminés du site actif. | Une enzyme peut perdre sa spécificité si le pH s’éloigne trop de sa zone optimale. La pepsine fonctionne vers pH 2, alors que beaucoup d’enzymes cytosoliques préfèrent un pH proche de 7. |
| Température | Elle augmente d’abord la fréquence des collisions, puis peut dénaturer la protéine. | À basse température, la réaction ralentit ; à trop haute température, la structure se défait. Chez l’humain, beaucoup d’enzymes travaillent au voisinage de 37 °C. |
| Concentration en substrat | Elle augmente la probabilité de formation du complexe jusqu’à saturation. | On observe une montée de vitesse, puis un plafond quand tous les sites actifs sont occupés. |
| Cofacteurs et ions métalliques | Ils peuvent stabiliser la forme active ou participer directement à la catalyse. | Sans eux, certaines enzymes restent incomplètes ou inactives. |
| Inhibiteurs et mutations | Ils perturbent la reconnaissance ou la géométrie du site actif. | Une petite modification peut réduire l’affinité, changer la vitesse ou, au contraire, ouvrir de nouvelles possibilités de liaison. |
Si l’on veut aller un peu plus loin, le paramètre Km est souvent utile : il donne une idée de la concentration de substrat nécessaire pour atteindre la moitié de la vitesse maximale. Je le considère comme un indice pratique, pas comme une vérité absolue, parce qu’il dépend du contexte expérimental et de la dynamique réelle de l’enzyme.
Ces variations ne décrivent pas seulement la chimie du moment. Elles expliquent aussi pourquoi les enzymes ont pu évoluer, se spécialiser ou parfois élargir leur champ d’action.
Pourquoi ce complexe est au cœur de l’évolution des enzymes
En biologie évolutive, une enzyme n’est jamais seulement une machine chimique : c’est aussi un objet façonné par la sélection naturelle. Le site actif, la flexibilité de la protéine et les charges électriques qui entourent la poche de liaison ont tous pu être affinés au fil du temps. Le complexe enzyme-substrat reflète donc une histoire d’adaptation, pas une simple coïncidence structurale.
Une spécificité forte, mais rarement absolue
On dit souvent qu’une enzyme est spécifique de son substrat, et c’est vrai dans l’ensemble. Mais cette spécificité n’est presque jamais absolue. Beaucoup d’enzymes acceptent plusieurs molécules proches, à condition que leur forme ou leur chimie reste compatible avec le site actif. C’est une contrainte, bien sûr, mais aussi une marge de manœuvre évolutive.
La promiscuité enzymatique comme point de départ
Une enzyme peut présenter une faible activité sur un substrat secondaire. Cette promiscuité enzymatique est loin d’être un défaut anecdotique : elle fournit souvent la matière première de l’innovation évolutive. Si une mutation améliore légèrement cette activité secondaire, la sélection peut ensuite amplifier ce gain au fil des générations.
Autrement dit, l’évolution ne repart pas toujours de zéro. Elle recycle ce qui existe déjà, puis ajuste la poche de liaison, la dynamique de la protéine ou la répartition des charges pour améliorer une interaction particulière.
Lire aussi : Réplication ADN - Le guide pour enfin comprendre les schémas
Mutation, duplication et sélection
Les mutations qui touchent le site actif peuvent avoir trois effets principaux : renforcer la liaison, la réduire ou changer le spectre des substrats reconnus. La duplication d’un gène enzymatique ajoute encore une possibilité intéressante : une copie conserve la fonction d’origine tandis que l’autre peut explorer de nouvelles variantes sans mettre en péril la fonction initiale.
C’est une logique très puissante. Chez certaines bactéries, par exemple, des enzymes capables d’hydrolyser des antibiotiques illustrent bien cette plasticité : une modification modeste du site actif peut suffire à élargir la gamme de molécules reconnues et à changer le sort d’une population entière face à un médicament.
Pour rendre cette logique plus concrète, quelques exemples simples parlent mieux qu’une définition abstraite.
Quelques exemples qui rendent la notion plus concrète
Quand j’explique ce sujet, je préfère toujours partir d’exemples connus. Ils montrent que le complexe n’est pas une abstraction de laboratoire, mais un mécanisme quotidien du vivant. On le retrouve dans la digestion, la détoxification cellulaire, la réplication de l’ADN et bien d’autres fonctions encore.
| Enzyme | Substrat | Ce que le complexe permet |
|---|---|---|
| Lactase | Lactose | Elle hydrolyse le sucre du lait en molécules plus simples, ce qui illustre très bien l’importance de la complémentarité de forme. |
| Amylase | Amidon | Elle amorce la digestion des polysaccharides et montre qu’un même type d’enzyme peut agir sur des chaînes longues si le site actif les positionne correctement. |
| Catalase | Peroxyde d’hydrogène | Elle accélère une réaction de détoxification très rapide, utile pour limiter l’accumulation de composés réactifs. |
| Lysozyme | Peptidoglycane bactérien | Il fragilise la paroi bactérienne en ciblant une structure précise, ce qui montre que la reconnaissance du substrat peut aussi reposer sur plusieurs contacts successifs. |
Ce tableau met en évidence une idée simple : ce n’est pas seulement la présence du substrat qui compte, mais la façon dont il s’insère dans le site actif et s’y aligne. Le complexe enzyme-substrat agit donc comme un dispositif d’orientation fine, plus que comme une simple colle moléculaire.
Le point à retenir pour lire une enzyme comme un système vivant
Quand j’analyse une enzyme, je ne regarde jamais seulement si elle fonctionne. Je regarde avec quel substrat elle fonctionne, dans quelles conditions, et quel compromis évolutif a pu conduire à cette forme de spécificité. C’est souvent là que l’on comprend vraiment la logique du vivant.
- Une forte affinité n’est pas toujours synonyme de meilleure catalyse.
- Une spécificité élevée peut être un avantage, mais aussi une contrainte.
- Une mutation utile dans un environnement peut devenir défavorable dans un autre.
- Le complexe enzyme-substrat résume bien cette réalité : efficacité, souplesse et limite font partie du même système.
Si je devais garder une seule idée, ce serait celle-ci : une enzyme n’est pas une pièce rigide, mais une structure dynamique qui reconnaît, ajuste et transforme. C’est précisément cette souplesse qui explique à la fois la puissance des réactions biologiques et la capacité des enzymes à évoluer.