L’hydrolyse de l’ATP est l’un des gestes chimiques les plus importants du vivant : elle permet à la cellule de convertir une réserve d’énergie en travail utile, qu’il s’agisse de transport membranaire, de mouvement, de synthèse ou de signalisation. Dans cet article, je vais aller droit au mécanisme, puis montrer pourquoi cette réaction est si efficace, comment les cellules l’exploitent au quotidien et ce qu’elle révèle sur l’évolution du vivant.
Les points essentiels à garder en tête
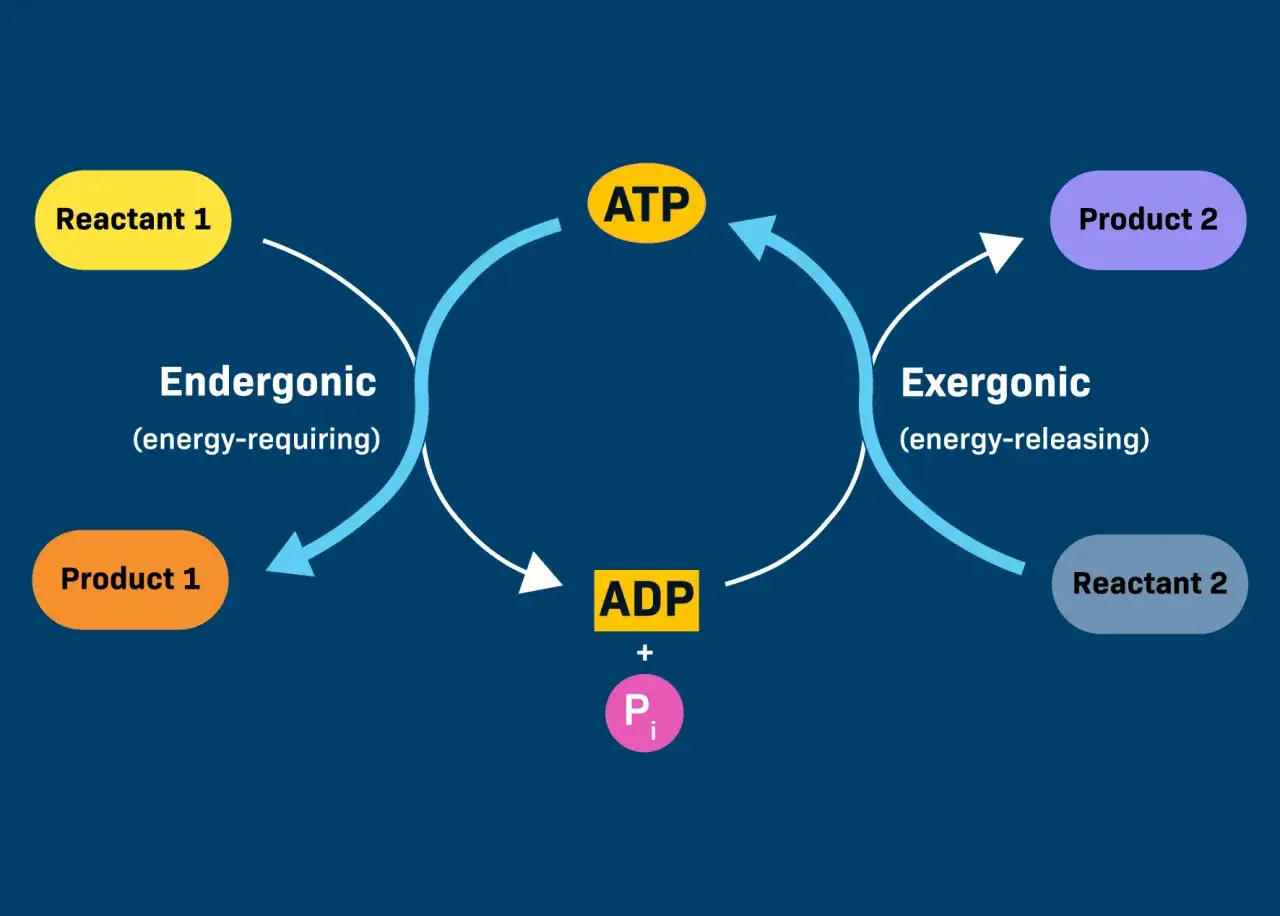
- ATP + H2O → ADP + phosphate inorganique est la réaction de base, mais l’intérêt biologique vient du couplage, pas d’une “énergie magique” libérée par simple cassure de liaison.
- En conditions biochimiques standard, le ΔG°’ est d’environ −30,5 kJ/mol; dans la cellule, la valeur réelle est souvent plus négative selon les concentrations.
- L’ATP sert surtout à alimenter le transport actif, la contraction, la biosynthèse et la phosphorylation.
- Le magnésium et les enzymes spécialisées rendent cette réaction exploitable et contrôlée.
- Sur le plan évolutif, l’ATP s’est imposé parce qu’il combine stabilité, réactivité et universalité.
Ce que fait réellement l’ATP quand il est hydrolysé
La réaction la plus classique s’écrit simplement : ATP + H2O → ADP + Pi. Le Pi désigne le phosphate inorganique, c’est-à-dire le groupement phosphate libéré après l’ajout d’une molécule d’eau. En pratique, la cellule coupe surtout le phosphate terminal, celui qui est le plus souvent mobilisé pour fournir un gain énergétique immédiatement exploitable.
Je préfère corriger tout de suite un raccourci trompeur : on dit souvent que l’ATP “contient de l’énergie dans ses liaisons”. C’est commode pour apprendre, mais insuffisant pour comprendre. Une liaison chimique ne libère pas de l’énergie par le seul fait d’être rompue ; la valeur biologique de cette réaction vient du bilan global entre les réactifs et les produits, ainsi que du contexte enzymatique.
Dans certains cas, l’ATP ne va pas jusqu’à l’ADP : il peut aussi être transformé en AMP et pyrophosphate (PPi), notamment lors de réactions de biosynthèse où il faut activer fortement une molécule. Cette variante est plus coûteuse, mais elle permet à la cellule de pousser des réactions autrement peu favorables.
Autrement dit, ce n’est pas une simple “vidange” d’énergie. C’est un système de transfert très précis, et c’est ce déplacement contrôlé du phosphate terminal qui prépare la vraie question : d’où vient l’énergie effectivement utilisable par la cellule ?
Pourquoi cette réaction libère une énergie exploitable
L’intérêt de cette hydrolyse vient de plusieurs effets qui se renforcent. D’abord, les produits ADP et Pi sont en général plus stables que l’ATP d’origine. Ensuite, les charges négatives des phosphates sont mieux réparties après la réaction. Enfin, l’environnement aqueux stabilise plus facilement les produits que la molécule de départ dans de nombreuses conditions cellulaires.
En conditions biochimiques standard, le changement d’énergie libre est d’environ −30,5 kJ/mol. Dans une cellule vivante, la valeur réelle dépend du rapport ATP/ADP/Pi, du pH, de la présence de magnésium et du compartiment concerné. Dans beaucoup de contextes cellulaires, la réaction devient donc encore plus favorable, ce qui explique qu’elle puisse financer des opérations exigeantes sans se gripper immédiatement.
Le point le plus important, à mon sens, est celui-ci : la cellule n’utilise pas l’ATP comme un carburant qui “explose”. Elle l’utilise comme un intermédiaire de couplage. Une réaction défavorable peut être rendue possible si elle est associée à une étape d’hydrolyse suffisamment favorable, à condition que l’enchaînement soit enzymatiquement bien orchestré.
Cette logique de couplage change tout. Une fois comprise, on voit pourquoi la même molécule peut alimenter des tâches très différentes sans que le système cellulaire perde sa cohérence.
Comment la cellule s’en sert pour produire du mouvement et du transport
L’ATP hydrolysé n’alimente pas une seule fonction : il sert à des dizaines de processus. Certains sont mécaniques, d’autres chimiques, d’autres encore concernent les gradients ioniques. Pour y voir clair, je résume les usages les plus parlants dans le tableau ci-dessous.
| Fonction cellulaire | Ce que l’ATP apporte | Exemple concret |
|---|---|---|
| Transport actif | Changement de conformation d’une pompe membranaire | La pompe Na+/K+ fait sortir 3 Na+ et entrer 2 K+ par ATP |
| Mouvement | Cycle mécanique d’un moteur moléculaire | La myosine sur l’actine pendant la contraction musculaire |
| Biosynthèse | Activation d’un substrat pour le rendre réactif | Synthèse des protéines, de l’ADN ou de certains lipides |
| Signalisation | Ajout ou retrait de groupements phosphate | Kinases et cascades de phosphorylation |
| Organisation cellulaire | Remodelage de structures protéiques | Assemblage et désassemblage du cytosquelette |
Ce tableau montre une idée essentielle : la cellule ne se contente pas de “consommer” de l’ATP, elle le convertit en transformations très ciblées. Le même principe vaut pour des systèmes aussi différents que la contraction d’un sarcomère, la mise en place d’un gradient de sodium ou l’activation d’un acide aminé avant incorporation dans une protéine.
Il y a aussi un effet de lecture métabolique. Quand le niveau d’ATP baisse et que l’ADP ou l’AMP augmentent, la cellule interprète cette variation comme un signal d’alerte énergétique. C’est l’une des raisons pour lesquelles l’ATP est bien plus qu’un simple “réservoir” : il participe à la régulation globale du fonctionnement cellulaire.
Reste à comprendre comment la cellule rend ce couplage possible avec suffisamment de finesse pour éviter le gaspillage et le chaos chimique.
Le magnésium et les enzymes qui rendent la réaction utile
Dans la cellule, l’ATP n’agit presque jamais seul. Il est souvent lié à un ion magnésium, sous la forme MgATP. Ce détail est fondamental : le magnésium aide à neutraliser une partie des charges négatives des phosphates et facilite la reconnaissance de l’ATP par les enzymes. Sans lui, beaucoup d’interactions seraient moins stables et moins efficaces.
Le rôle du magnésium
Le magnésium ne “crée” pas l’énergie, mais il rend la chimie plus maniable. Il aide à orienter les phosphates, à réduire les répulsions électrostatiques et à stabiliser l’état de liaison dans les sites actifs. En clair, il transforme une molécule très chargée en substrat exploitable par des protéines spécialisées.
Lire aussi : Hardy-Weinberg - Comprendre l'équilibre et ses écarts
ATPases et kinases ne font pas la même chose
On confond souvent les deux. Les ATPases hydrolysent l’ATP pour provoquer un changement de forme, un déplacement ou un transport. Les kinases, elles, transfèrent généralement un phosphate de l’ATP vers une autre molécule. La première catégorie dépense de l’énergie pour produire un effet mécanique ou conformationnel ; la seconde modifie l’état chimique d’une cible.
Cette distinction semble technique, mais elle évite beaucoup de contresens. Quand une kinase phosphoryle une protéine, elle ne “brûle” pas simplement de l’ATP : elle reprogramme la protéine, parfois de manière réversible, pour modifier son activité, sa localisation ou ses partenaires.
Pourquoi l’ATP s’est imposé au cours de l’évolution
Si l’on regarde l’hydrolyse de l’ATP sous un angle évolutif, une question intéressante apparaît : pourquoi cette molécule-là est-elle devenue la monnaie énergétique dominante, et pas une autre ? La réponse tient à un équilibre très rare entre disponibilité chimique, stabilité et polyvalence. L’ATP est assez réactif pour être utile, mais pas au point de se dégrader trop vite en solution. Il est assez universel pour être reconnu par d’innombrables enzymes, mais assez simple pour être recyclé en permanence.
L’ATP a aussi bénéficié d’un effet de réseau. Une fois qu’un grand nombre d’enzymes, de pompes et de moteurs ont été construits autour de lui, le remplacer devient extraordinairement coûteux pour un organisme. L’évolution n’optimise pas à neuf à chaque génération ; elle bricole à partir de ce qui existe déjà. C’est pour cela qu’une solution ancienne, efficace et bien intégrée a de fortes chances de persister.
La présence quasi universelle de l’ATP synthase dans les êtres vivants va dans ce sens. Cette machine moléculaire est au cœur de la production d’ATP à partir d’un gradient de protons. Son ancienneté suggère que le couplage entre membrane, gradient chimique et synthèse d’ATP s’est installé très tôt dans l’histoire du vivant. Je reste prudent sur les détails exacts de cette émergence, car les scénarios de l’origine de la vie continuent d’évoluer, mais la tendance générale est claire : l’ATP n’est pas une invention tardive, c’est un pilier ancien de la bioénergétique.Cette robustesse évolutive éclaire aussi la raison pour laquelle le vivant a conservé l’ATP au lieu de le remplacer par une autre monnaie unique.
Les erreurs de compréhension qui reviennent sans cesse
- “L’énergie vient de la rupture de la liaison” : formulé ainsi, c’est incomplet. Le bilan énergétique dépend du système entier, pas de la seule cassure.
- “L’ATP est une batterie” : l’image aide au départ, mais elle masque le fait que l’ATP est surtout un intermédiaire recyclé en continu.
- “Toutes les réactions à l’ATP se ressemblent” : non. Certaines déplacent une pompe, d’autres phosphorylent une protéine, d’autres activent un substrat.
- “Le contexte n’a pas d’importance” : au contraire, le pH, les concentrations et les ions présents changent fortement l’énergie réellement disponible.
Je trouve ce point important parce qu’il sépare une compréhension scolaire d’une compréhension réellement biologique. Dans la cellule, la chimie n’est jamais isolée : elle est toujours enchâssée dans un environnement, des enzymes et des contraintes évolutives.
Ce qu’il faut garder en tête pour lire la biochimie cellulaire
Si je devais résumer l’essentiel en trois idées, je dirais ceci : l’ATP n’est pas une source d’énergie brute, c’est un système de couplage ; la cellule exploite surtout des réactions enzymatiques contrôlées, pas une simple cassure de liaison ; et l’évolution a retenu l’ATP parce qu’il était déjà très bon dans ce rôle.
Cette manière de voir change la lecture de toute la biologie cellulaire. Dès qu’une cellule transporte des ions, contracte un muscle, assemble une protéine ou active un substrat, il y a souvent derrière la même logique fondamentale : une hydrolyse bien placée, une enzyme qui la canalise et un besoin biologique précis qui en tire parti. C’est là que l’ATP cesse d’être une formule de manuel et devient une véritable clé de compréhension du vivant.
Si vous continuez à explorer la bioénergétique, le meilleur prolongement naturel est d’observer comment l’ATP synthase fabrique cette molécule à partir des gradients de protons, puis comment les cellules arbitrent en permanence entre production, consommation et signal de stress énergétique.