
Les protéines ne sont pas des objets opaques: elles se construisent à partir de briques chimiques simples, et c’est là que la biologie devient vraiment lisible. Je vais montrer comment les acides aminés s’assemblent, pourquoi leur architecture dicte la forme d’une protéine, et comment l’évolution a fixé un noyau commun chez presque tous les êtres vivants. Vous verrez aussi ce que cette logique change pour l’humain, notamment dans l’alimentation et la compréhension des mutations.
Les repères essentiels à garder sur ces briques du vivant
- Vingt formes standard servent de base à la plupart des protéines, avec quelques exceptions rares.
- Le groupe R, ou chaîne latérale, décide du comportement chimique de chaque unité.
- La liaison peptidique transforme une suite de briques en chaîne polypeptidique.
- Chez l’adulte, 9 de ces molécules doivent venir de l’alimentation.
- Le code génétique est presque universel, ce qui relie toute la vie à une histoire commune.
Ce que ces molécules apportent à la chimie du vivant
Je les vois comme des molécules-pivot: un carbone central, un groupe amine, un groupe carboxyle et une chaîne latérale, le groupe R. Cette architecture est simple sur le papier, mais elle permet une diversité énorme, parce que le R peut être hydrophobe, chargé, aromatique, soufré ou capable de former des liaisons faibles très sélectives. À pH physiologique, beaucoup adoptent une forme zwitterion, ce qui explique leur sensibilité au milieu, aux enzymes et aux variations de charge.
Deux cas sont particulièrement parlants. La glycine, avec une chaîne latérale réduite à un hydrogène, apporte de la souplesse; la proline, au contraire, rigidifie le squelette et crée souvent des coudes dans la chaîne. C’est exactement le genre de détail qui fait basculer une protéine d’une forme à une autre. Cette chimie de départ prend tout son sens quand la cellule commence à les relier en chaîne.
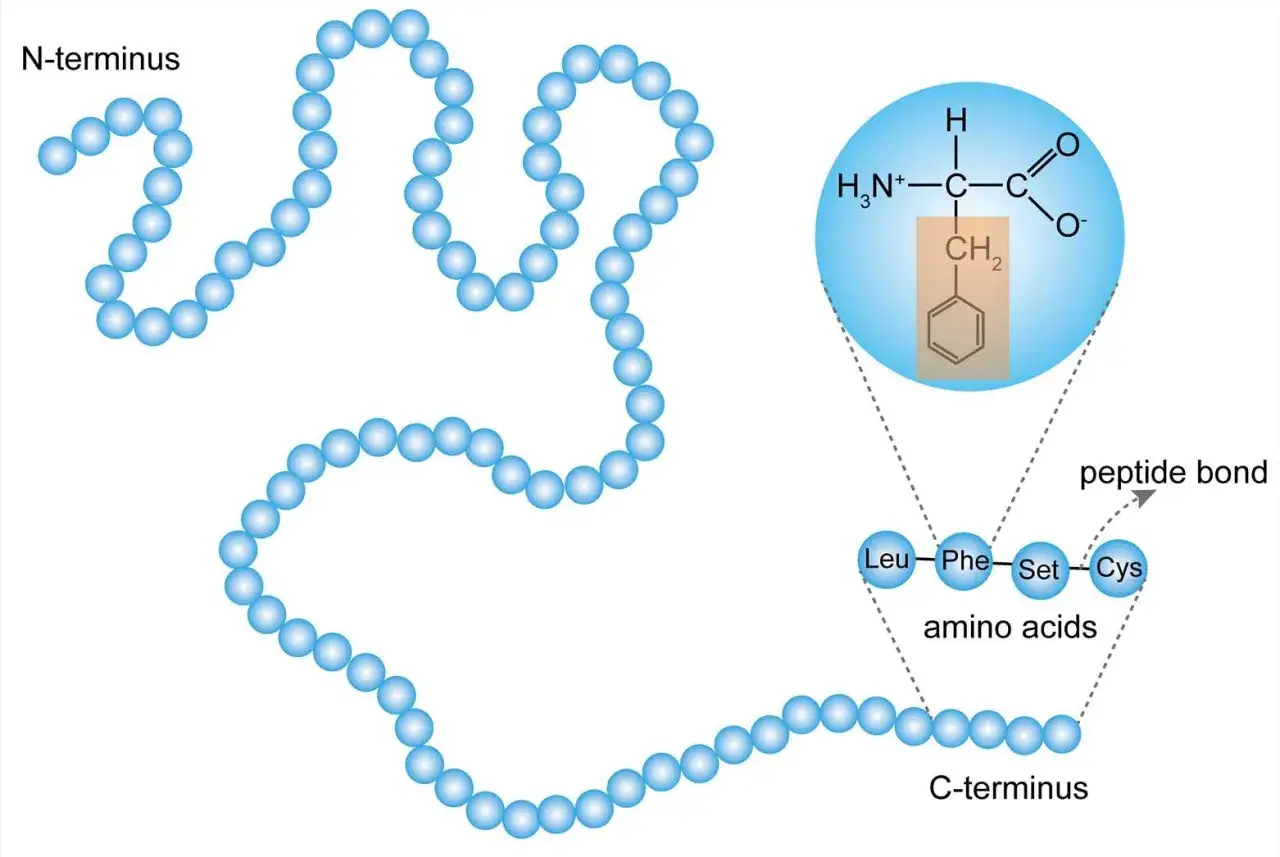
Comment une chaîne peptidique devient une protéine
Dans la cellule, le ribosome lit l’ARN messager et assemble les briques une à une par liaison peptidique, une condensation qui libère une molécule d’eau à chaque ajout. Une courte chaîne est souvent décrite comme un peptide; au-delà, on parle de polypeptide ou de protéine selon le contexte. Ce qui compte, ce n’est pas seulement la longueur, mais l’ordre précis des unités: une permutation minuscule peut modifier le repliement, l’activité enzymatique ou la stabilité thermique.
Je trouve utile de penser à ce moment comme à une traduction, pas comme à un simple collage. Le ribosome ne fabrique pas une matière brute: il convertit une information génétique en architecture tridimensionnelle, et les chaperonnes moléculaires aident parfois la chaîne à adopter la bonne forme. Sans repliement correct, une séquence prometteuse reste souvent biologiquement muette. Les biochimistes parlent ici de structure primaire, puis de structures plus complexes quand la chaîne commence à se plier et à se stabiliser.
Une fois la chaîne formée, il faut regarder ce que chaque famille chimique permet vraiment dans l’espace.
Les familles chimiques qui changent tout
Les propriétés des chaînes latérales expliquent pourquoi certaines portions d’une protéine s’enfouissent au cœur de la structure alors que d’autres restent exposées à l’eau. Pour lire une protéine, je commence presque toujours par là. Le tableau ci-dessous résume les familles les plus utiles à retenir.
| Famille | Exemples | Ce que cela change |
|---|---|---|
| Hydrophobes | Leucine, isoleucine, valine | Ils s’enterrent souvent au cœur des protéines et stabilisent le repliement. |
| Polaires non chargés | Sérine, thréonine, asparagine, glutamine | Ils forment facilement des liaisons hydrogène et restent volontiers en surface. |
| Acides | Aspartate, glutamate | Ils portent une charge négative et participent à la catalyse et aux interactions ioniques. |
| Basiques | Lysine, arginine, histidine | Ils portent une charge positive ou variable, utile pour l’ADN et les sites actifs. |
| Soufrés | Cystéine, méthionine | Ils peuvent former des ponts disulfure et jouer un rôle clé dans l’architecture des protéines. |
| Aromatiques | Phénylalanine, tyrosine, tryptophane | Ils participent aux empilements, à la reconnaissance moléculaire et à l’absorption UV. |
La glycine et la proline méritent un regard à part: l’une apporte de la souplesse, l’autre impose des angles serrés. Je m’en sers souvent comme repères rapides, parce qu’ils expliquent à eux seuls pourquoi certaines zones d’une protéine sont mobiles et d’autres très verrouillées. La cystéine, elle, peut former des ponts disulfure qui rigidifient fortement certaines structures, tandis que l’histidine change facilement d’état de charge et devient précieuse dans les sites actifs des enzymes.
Ce classement n’est pas seulement scolaire: il prédit la forme, la stabilité et, au bout du compte, la fonction. C’est précisément cette cohérence chimie-fonction que l’évolution a conservée.
Pourquoi l’évolution a stabilisé ce vocabulaire
Le point fascinant, c’est que ce vocabulaire est presque universel. D’une bactérie à l’humain, le code génétique associe presque toujours les mêmes codons aux mêmes briques, ce qui suggère une origine très ancienne et une forte contrainte évolutive. L’histoire du vivant n’a donc pas inventé chaque protéine de zéro; elle a travaillé avec un alphabet déjà installé, en le remaniant sans cesse.
Je trouve aussi très parlant que la structure 3D soit souvent mieux conservée que la séquence elle-même. En d’autres termes, l’évolution tolère certains remplacements tant que le repliement, la stabilité ou le site actif restent compatibles avec la fonction. C’est pour cela qu’une protéine peut changer de visage sans cesser de faire son travail.
Les exceptions rares, comme la sélénocystéine, rappellent malgré tout que ce système n’est pas figé; il peut s’étendre, mais seulement dans des cadres très précis. Au fond, cela raconte une règle simple: la vie innove, mais elle n’efface pas le socle sur lequel elle a commencé. Cette logique devient très concrète dès qu’on se demande ce que cela implique pour l’humain.
Ce que cela implique pour l’humain et son alimentation
Chez l’adulte, 9 de ces molécules sont dites indispensables: histidine, isoleucine, leucine, lysine, méthionine, phénylalanine, thréonine, tryptophane et valine. La liste peut varier un peu selon l’âge ou l’état physiologique, ce qui me semble important à rappeler, parce que la biologie aime les nuances plus que les slogans. “Non essentiel” ne veut pas dire secondaire: cela signifie simplement que l’organisme peut le synthétiser à partir d’autres molécules.
- Les protéines animales et végétales n’apportent pas exactement le même profil, donc la variété compte davantage qu’une seule source répétée.
- La quantité brute ne suffit pas toujours: la digestibilité et la composition réelle de l’apport font une vraie différence.
- Un excès d’un seul composant isolé compense rarement un régime pauvre et déséquilibré.
- En croissance, pendant la grossesse, à la récupération ou avec l’âge, les besoins peuvent changer sans que la logique de base change.
En pratique, ce que je conseille surtout de retenir, c’est qu’une stratégie alimentaire sérieuse regarde l’équilibre global des sources protéiques, pas seulement les grammes affichés. Cette lecture plus fine aide à éviter deux erreurs fréquentes: croire qu’une source unique suffit toujours, ou supposer qu’un complément isolé résout tout. Une fois ces repères en tête, on lit la nutrition avec le même œil analytique que la biologie moléculaire.
Lire la biologie avec cet alphabet en tête
Quand on regarde une mutation, une comparaison d’espèces ou un écart de digestion, la bonne question n’est pas seulement “quelle unité a changé ?”, mais “qu’a fait ce changement à la charge, à la taille, à la rigidité ou au repliement ?”. C’est là que la chimie rejoint l’histoire du vivant. Une substitution apparemment minime peut déplacer une interaction, perturber une enzyme ou, au contraire, améliorer une adaptation.
Je trouve que c’est la meilleure manière de lire les protéines: non pas comme des listes de lettres, mais comme des objets vivants, hérités et bricolés par l’évolution. À partir de ce niveau de lecture, le rôle des briques moléculaires devient beaucoup plus clair, et tout le reste de la biologie gagne en cohérence.