
Je vais donc partir de la structure de base, passer par la charge en solution, puis montrer comment ces petites molécules racontent aussi quelque chose de l’évolution: pourquoi le vivant a retenu certaines formes, comment il en tolère d’autres, et ce que cela change dans les protéines.
Les repères essentiels à garder en tête
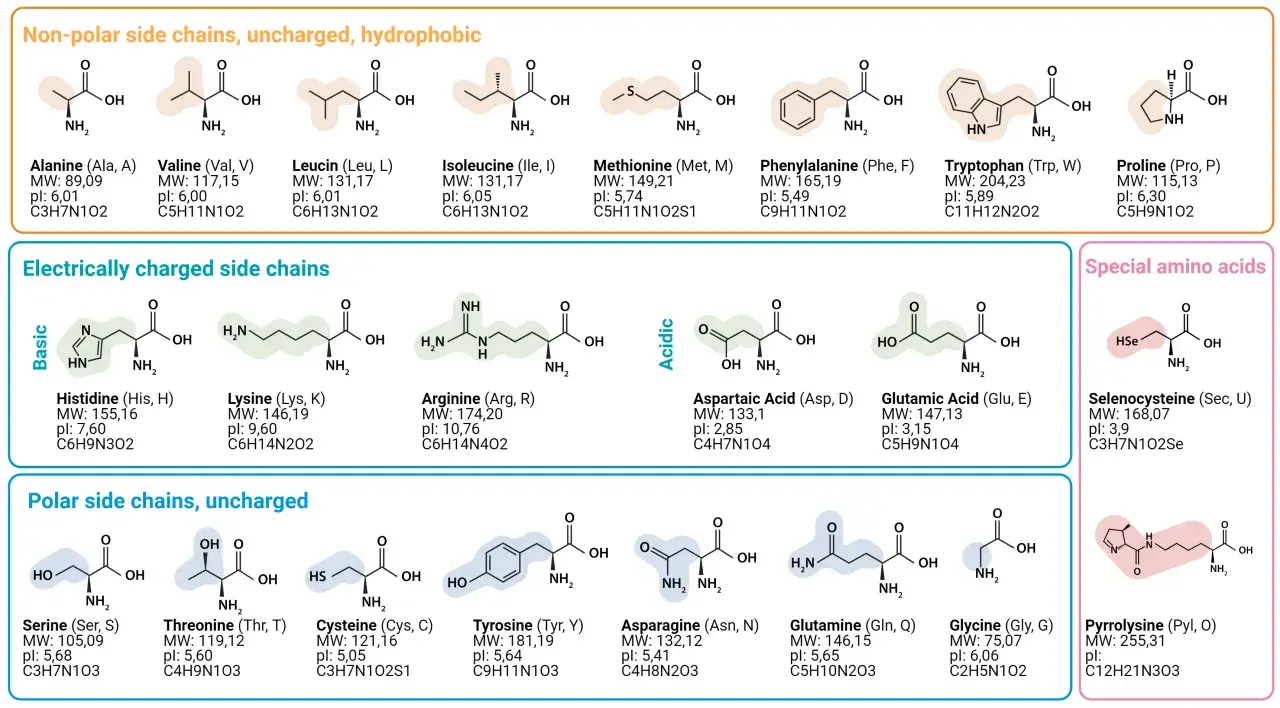
- Un acide aminé protéinogène repose sur un carbone alpha portant un groupe amine, un groupe carboxyle, un hydrogène et une chaîne latérale R.
- La chaîne R détermine la polarité, la charge, la taille et une grande partie du comportement biologique.
- En solution aqueuse, beaucoup d’acides aminés existent sous forme zwitterionique, avec une charge positive et une charge négative sur la même molécule.
- Les protéines sont construites à partir de 20 acides aminés standard, avec quelques exceptions rares dans certains organismes.
- L’évolution conserve surtout ce qui préserve le repliement, l’activité et l’économie énergétique.
Le squelette commun des acides aminés
Le point de départ est toujours le même: un carbone alpha central auquel sont liés quatre éléments. On trouve un groupe amine, un groupe carboxyle, un atome d’hydrogène et une chaîne latérale, souvent notée R. Cette organisation explique pourquoi les acides aminés forment une famille cohérente tout en restant chimiquement très différents les uns des autres.
| Élément | Rôle dans la molécule | Ce qu’il faut retenir |
|---|---|---|
| Carbone alpha | Centre de la structure | Il relie les quatre groupes et crée l’architecture commune. |
| Groupe amine | Comportement basique | Il peut capter un proton et participer à la liaison peptidique. |
| Groupe carboxyle | Comportement acide | Il peut céder un proton et former la base des liaisons peptidiques. |
| Hydrogène | Complète la géométrie du carbone alpha | Il participe à l’asymétrie de la molécule. |
| Chaîne latérale R | Variable | Elle détermine l’essentiel des propriétés biologiques. |
Deux cas méritent d’être mémorisés tout de suite. La glycine a pour chaîne latérale un simple hydrogène, ce qui la rend achirale et particulièrement souple. La proline, elle, ferme sa chaîne latérale sur l’azote du squelette, ce qui rigidifie la molécule et influence fortement le repliement des protéines.
Autrement dit, le squelette est commun, mais il ne dit pas encore comment la molécule va se comporter. Pour cela, il faut regarder la chaîne R de près.
La chaîne latérale explique la vraie diversité
C’est la chaîne latérale qui transforme une famille chimique uniforme en palette fonctionnelle. Selon sa taille, sa polarité ou sa charge, un acide aminé va préférer l’eau, fuir l’eau, stabiliser une structure ou intervenir dans une réaction enzymatique. Je trouve utile de penser cette chaîne R comme la partie « signature » de la molécule.
| Classe | Exemples | Effet principal |
|---|---|---|
| Non polaires, hydrophobes | Valine, leucine, isoleucine, phénylalanine, méthionine | Ils se cachent souvent au cœur des protéines et favorisent un noyau compact. |
| Polaires non chargés | Sérine, thréonine, asparagine, glutamine | Ils forment facilement des liaisons hydrogène et sont fréquents à la surface. |
| Acides | Aspartate, glutamate | Ils portent une charge négative et participent aux ponts salins et à la catalyse. |
| Basiques | Lysine, arginine, histidine | Ils portent une charge positive, utile pour interagir avec l’ADN et d’autres molécules chargées. |
Cette classification est pratique, mais elle n’est pas figée. Tyrosine, cystéine ou histidine peuvent changer de comportement selon le contexte, le pH ou l’environnement local de la protéine. C’est précisément ce genre de nuance qui fait la richesse de la biochimie: une même formule ne raconte pas tout, il faut aussi regarder le milieu.
Et c’est là que la charge des acides aminés devient décisive, car en solution aqueuse ils ne gardent pas toujours la même forme.
Pourquoi le pH change leur charge
En eau, les acides aminés sont souvent présents sous forme zwitterionique, c’est-à-dire avec une charge positive et une charge négative sur la même molécule. Le groupe carboxyle peut perdre un proton, tandis que le groupe amine peut en capter un. Résultat: à pH proche du neutre, la molécule se comporte rarement comme un composé strictement neutre.
Le point isoélectrique est le pH auquel la charge nette d’un acide aminé est nulle. Il ne faut pas le confondre avec les valeurs où les groupes fonctionnels basculent eux-mêmes: autour de pH 2,2, le carboxyle change nettement de comportement, et autour de pH 9,4, c’est l’amine qui bascule à son tour. Ces seuils varient un peu selon la chaîne R, donc je préfère les voir comme des repères utiles plutôt que comme des constantes absolues.
| Milieu | Forme dominante | Conséquence fréquente |
|---|---|---|
| Très acide, autour de pH 2,2 ou moins | Forme davantage protonée | La charge globale tend vers le positif. |
| Proche du point isoélectrique | Zwitterion majoritaire | La charge nette est proche de zéro, la solubilité peut diminuer. |
| Basique, autour de pH 9,4 ou plus | Forme davantage déprotonée | La charge globale tend vers le négatif. |
Cette logique explique pourquoi le comportement d’un acide aminé en laboratoire ou dans une cellule dépend autant du milieu que de sa formule brute. Une fois ce mécanisme compris, la liaison entre acides aminés et protéines devient beaucoup plus lisible.
De l’acide aminé à la protéine
Un acide aminé devient vraiment intéressant lorsqu’il s’insère dans une chaîne. Le carbone du groupe carboxyle d’un acide aminé se lie à l’azote du groupe amine d’un autre: c’est la liaison peptidique. Cette condensation libère une molécule d’eau et crée une ossature répétitive, le polypeptide.
| Partie de la protéine | Nature | Rôle biologique |
|---|---|---|
| Ossature peptidique | Répétitive et relativement rigide | Elle porte les motifs structuraux de base, comme l’hélice alpha et le feuillet beta. |
| Chaînes latérales R | Variables | Elle commandent les interactions, la reconnaissance moléculaire et la spécificité fonctionnelle. |
La liaison peptidique est presque plane et ne tourne pas librement, ce qui limite la géométrie possible de la chaîne. En revanche, les chaînes R orientent le repliement: les groupes hydrophobes se cachent souvent au centre, tandis que les groupes polaires se placent plus volontiers en surface, là où ils peuvent interagir avec l’eau.
C’est aussi la raison pour laquelle une seule substitution peut parfois tout changer. Remplacer un résidu hydrophobe par un résidu chargé, ou l’inverse, peut modifier la stabilité, le site actif ou la forme finale de la protéine. Pour comprendre pourquoi le vivant a retenu précisément ces acides aminés et pas d’autres, il faut passer à la lecture évolutive.
Ce que l’évolution a retenu
Sur le plan évolutif, les acides aminés ne sont pas apparus comme une liste arbitraire de vingt noms. Le code génétique standard repose sur 20 acides aminés utilisés dans la synthèse ribosomique, avec quelques cas particuliers rares comme la sélénocystéine et la pyrrolysine dans certains organismes. J’aime retenir une idée simple: le vivant a gardé un petit alphabet chimique parce qu’il était suffisamment riche pour construire des protéines efficaces, mais pas trop complexe pour rester gérable.
La homochiralité est un autre point clé. Les protéines sont presque exclusivement construites avec des L-acides aminés, à l’exception de la glycine, qui n’est pas chirale. Cette uniformité de « main » moléculaire facilite le repliement correct; un mélange désordonné de formes L et D brouillerait la géométrie de la chaîne. Les D-acides aminés existent tout de même dans la nature, notamment dans certaines parois bactériennes et dans quelques peptides spécialisés.
- La séquence compte, mais la structure compte encore plus. Des protéines homologues peuvent garder une architecture proche même avec moins de 30 % d’identité de séquence.
- Les substitutions conservatrices dominent. L’évolution remplace souvent un acide aminé par un autre de taille, de charge ou de propension structurale voisine.
- Le coût biosynthétique pèse dans la balance. Certains résidus sont plus « chers » à fabriquer, et cette contrainte influence leur fréquence au cours de l’évolution.
- La stabilité du repliement agit comme un filtre. Les positions du cœur protéique et du site actif tolèrent moins bien les changements que les régions de surface.
Autrement dit, l’évolution ne sélectionne pas seulement des fonctions: elle sélectionne aussi des architectures supportables. C’est pour cela que la chimie des chaînes R et l’histoire des protéines sont si étroitement liées.
Les repères qui évitent les confusions en schéma
Quand je lis un schéma d’acide aminé, je commence toujours par le même trio: le carbone alpha, la chaîne R et l’état de protonation. C’est le moyen le plus rapide de passer d’une formule un peu sèche à une lecture biologique utile.
- Repérer d’abord le carbone alpha, car c’est lui qui relie les quatre éléments fondamentaux.
- Vérifier si la chaîne latérale est hydrophobe, polaire, acide ou basique, car cela annonce déjà son comportement.
- Ne pas oublier que la charge dépend du pH et qu’elle n’est pas fixée une fois pour toutes.
- Se souvenir que glycine est achirale et que proline rigidifie la chaîne.
- Ne pas confondre l’alphabet standard des protéines avec tous les acides aminés possibles dans la nature.
Au fond, tout le sujet tient dans cette idée: une architecture minuscule, quelques variantes chimiques bien choisies, et une immense diversité de fonctions. C’est pour cela que comprendre la structure d’un acide aminé revient déjà à entrer dans la logique de la biologie et de l’évolution des protéines.